

Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...

Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰)...
Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...
Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰)...
Топ:
Устройство и оснащение процедурного кабинета: Решающая роль в обеспечении правильного лечения пациентов отводится процедурной медсестре...
Техника безопасности при работе на пароконвектомате: К обслуживанию пароконвектомата допускаются лица, прошедшие технический минимум по эксплуатации оборудования...
Генеалогическое древо Султанов Османской империи: Османские правители, вначале, будучи еще бейлербеями Анатолии, женились на дочерях византийских императоров...
Интересное:
Лечение прогрессирующих форм рака: Одним из наиболее важных достижений экспериментальной химиотерапии опухолей, начатой в 60-х и реализованной в 70-х годах, является...
Мероприятия для защиты от морозного пучения грунтов: Инженерная защита от морозного (криогенного) пучения грунтов необходима для легких малоэтажных зданий и других сооружений...
Искусственное повышение поверхности территории: Варианты искусственного повышения поверхности территории необходимо выбирать на основе анализа следующих характеристик защищаемой территории...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
С деятельностью поверхностных рецепторов клеток связан такой феномен, как клеточная адгезия.
Адгезия — процесс взаимодействия специфических гликопротеинов соприкасающихся плазматических мембран распознающих друг друга клеток или клеток и внеклеточного матрикса. В том случае, если гликоиротеины при этом образуют связи, происходит адгезия, а затем формирование прочных межклеточных контактов или контактов клетки и межклеточного матрикса.
Все молекулы клеточной адгезии подразделяются на 5 классов.
1. Кадгерины. Это трансмембранные гликопротеины, использующие для адгезии ионы кальция. Отвечают за организацию цитоскелета, взаимодействие клеток с другими клетками.
2. Интегрины. Как уже отмечалось, интегрины представляют собой мембранные рецепторы для белковых молекул внеклеточного матрикса — фибронектина, ламинина и др. Связывают внеклеточный матрикс с цитос-келетом при помощи внутриклеточных белков талина, винкулина, а-акти-нина. Функционируют как клеточно-виеклеточные, так и межклеточные адгезионные молекулы.
3. Селектины. Обеспечивают прилипание лейкоцитов к эндотелию сосудов и тем самым — лейкоцитарно-эндотелиальпые взаимодействия, миграцию лейкоцитов через стенки сосудов в ткани.
4. Семейство иммуноглобулинов. Эти молекулы играют важную роль в иммунном ответе, а также в эмбриогенезе, заживлении ран и др.
5. Гоминговые молекулы. Обеспечивают взаимодействие лимфоцитов с эндотелием, их миграцию и заселение специфических зон иммунокомпс-тептных органов.
Таким образом, адгезия является важным звеном клеточной рецепции, играет большую роль в межклеточных взаимодействиях и взаимодействиях клеток с внеклеточным матриксом. Адгезионные процессы абсолютно необходимы при таких общебиологических процессах, как эмбриогенез, иммунный ответ, рост, регенерация и др. Они участвуют также в регуляции внутриклеточного и тканевого гомеостаза.
|
|
ЦИТОПЛАЗМА
ГИАЛОПЛАЗМА. Гиалоплазму называют также клеточным соком, цитозолем, или клеточным матриксом. Это основная часть цитоплазмы, составляющая около 55% объема клетки. В ней осуществляются основные клеточные обменные процессы. Гиалонлазма является сложной коллоидной системой и состоит из гомогенного мелкозернистого вещества с низ-кой электронной плотностью. Она состоит из воды, белков, нуклеиновых кислот, полисахаридов, липидов, неорганических веществ. Гиалоплазма может менять свое агрегатное состояние: переходить из состояния жидкого (золь) в более плотное — гель. При этом может изменяться форма клетки, ее подвижность и обмен веществ. Функции гиалонлазмы:
1. Метаболическая — метаболизм жиров, белков, углеводов.
2. Формирование жидкой микросреды (матрикса клетки).
3. Участие в движении клетки, обмене веществ и энергии. ОРГАНЕЛЛЫ. Органеллы — это второй важнейший обязательный
компонент клетки. Важным признаком органелл является то, что они имеют постоянное строго определенное строение и функции. По функциональному признаку все органеллы делятся на 2 группы:
1. Органеллы общего значения. Содержатся во всех клетках, поскольку необходимы для их жизнедеятельности. Такими органеллами являются: митохондрии, эндоплазматическая сеть (ЭПС) двух видов, комплекс Голь-джи (КГ), центриоли, рибосомы, лизосомы, пероксисомы, микротрубочки и микрофиламенты.
2. Органеллы специального значения. Есть только в тех клетках, которые выполняют специальные функции. Такими органеллами являются миофибриллы в мышечных волокнах и клетках, нейрофибриллы в нейронах, жгутики и реснички.
По структурному признаку все органеллы делятся на: 1) органеллы мембранного типа и 2) органеллы немембранного типа. Кроме того, немембранные органеллы могут быть построены по фибриллярному и гранулярному принципу.
|
|
В органеллах мембранного типа основным компонентом являются внутриклеточные мембраны. К таким органеллам относятся митохондрии, ЭПС, КГ, лизосомы, пероксисомы. К немембранным органеллам фибриллярного типа относятся микротрубочки, микрофиламенты, реснички, жгутики, центриоли. К немембранным гранулярным органеллам относят рибосомы, полисомы.
МЕМБРАННЫЕ ОРГАНЕЛЛЫ
ЭНДОПЛАЗМАТИЧЕСКАЯ СЕТЬ (ЭПС) - мембранная органелла, описанная в 1945 году К. Портером. Ее описание стало возможно благодаря электронному микроскопу. ЭПС — это система мелких каналов, вакуолей, мешочков, образующих в клетке непрерывную сложную сеть, элементы которой часто могут формировать кажущиеся на ультратонких срезах изолированными вакуоли. ЭПС построена из мембран, более тонких, чем цитолемма, и содержащих больше белка из-за находящихся в ней многочисленных ферментных систем. Есть 2 вида ЭПС: гранулярная (шероховатая) и агранулярная, или гладкая. Оба вида ЭПС могут взаимно переходить друг в друга и функционально связаны между собой так называемой переходной, или транзиторной, зоной.
Гранулярная ЭПС (рис. 3.3) содержит на своей поверхности рибосомы (полисомы) и является органеллой биосинтеза белка. Полисомы или рибосомы связываются с ЭПС при помощи так называемого причального белка (docking protein). При этом в мембране ЭПС имеются специальные интегральные белки рибофорины, также связывающие рибосомы и формирующие гидрофобные трапемембранные каналы для транспорта синтезированной полипентидной цени в просвет гранулярной ЭПС.
Гранулярная ЭПС видна только в электронном микроскопе. В световом микроскопе признаком развитой гранулярной ЭПС служит базофилия цитоплазмы. Гранулярная ЭПС имеется в каждой клетке, но степень ее развития различна. Она максимально развита в клетках, синтезирующих белок на экспорт, т.е. в секреторных клетках. Максимального развития гранулярная ЭПС достигает в нейроцитах, в которых ее цистерны приобретают упорядоченное расположение. В этом случае на светомикроскопическом уровне она выявляется в виде закономерно расположенных участков базофилии цитоплазмы, называемых базофилыюй субстанцией Ниссля.
|
|

|
Функция гранулярной ЭПС — синтез белка на экспорт. Кроме того, в ней происходят начальные посттрансляционные изменения полипептидной цепочки: гидроксилирование, сульфатирование и фосфорилиро-вание, гликозилирование. Последняя реакция особенно важна, т.к. приводит к образованию гликопротеинов — наиболее частого продукта клеточной секреции.
Агранулярная (гладкая) ЭПС представляет собой трехмерную сеть канальцев, не содержащих рибосомы. Гранулярная ЭПС может без перерыва переходить в гладкую ЭПС, но может существовать как самостоятельная органелла. Место перехода гранулярной ЭПС в агранулярную называется переходной (промежуточной, транзиторной) частью. От нее происходит отделение пузырьков с синтезированным белком и транспорт их к комплексу Гольджи.
Функции гладкой ЭПС:
1. Разделение цитоплазмы клетки на отделы — компартменты, в каждом из которых идет своя группа биохимических реакций.
2. Биосинтез жиров, углеводов.
3. Образование пероксисом;
4. Биосинтез стероидных гормонов;
5. Дезинтоксикация экзо- и эндогенных ядов, гормонов, биогенных аминов, лекарств за счет деятельности специальных ферментов.
6. Депонирование ионов кальция (в мышечных волокнах и миоцитах);
7. Источник мембран для восстановления кариолеммы в телофазе митоза.
ПЛАСТИНЧАТЫЙ КОМПЛЕКС ГОЛЬДЖИ. Это мембранная органелла, описанная в 1898 г. итальянским нейрогистологом К.Гольджи. Он назвал эту органеллу внутриклеточным сетчатым аппаратом из-за того, что в световом микроскопе она имеет сетчатый вид (рис. 3.4, а). Световая микроскопия не дает полного представления о строении этой органеллы. В световом микроскопе комплекс Гольджи имеет вид сложной сети, в которой ячейки могут быть связаны друг с другом или лежать независимо друг от друга (диктиосомы) в виде отдельных темных участков, палочек, зерен, вогнутых дисков. Между сетчатой и диффузной формой комплекса Гольд-жи нет принципиального различия, может наблюдаться смена форм этой оргамеллы. Еще в эпоху световой микроскопии было отмечено, что морфология комплекса Гольджи зависит от стадии секреторного цикла. Это позволило Д.Н.Насонову предположить, что комплекс Гольджи обеспечивает накопление синтезируемых веществ в клетке. По данным электронной микроскопии, комплекс Гольджи состоит из мембранных структур: плоских мембранных мешков с ампулярными расширениями на концах, а также крупных и мелких вакуолей (рис. 3.4, б, в). Совокупность этих образований называют диктиосомой. В диктиосоме находятся 5—10 мешковидных цистерн. Число диктиосом в клетке может достигать нескольких десятков. При этом каждая диктиосома связана с соседней при помощи вакуолей. В каждой диктиосоме есть проксимальная, незрелая, формирующаяся, или ЦИС-зона, -повернутая к ядру, и дистальная, ТРАНС-зона. Последняя, в отличие от выпуклой цис-поверхности, вогнутая, зрелая, обращена к цитолем- ме клетки. С цис-стороны происходит присоединение пузырьков, отделяющихся от переходной зоны ЭПС и содержащих вновь синтезированный и частично процессированный белок. При этом мембраны пузырьков встраиваются в мембрану цис-поверхности. С транс-стороны отделяются секреторные пузырьки и лизосомы. Таким образом, в комплексе Гольджи существует постоянный поток клеточных мембран и их созревание. Функции комплекса Гольджи:
|
|
1. Накопление, созревание и конденсация продуктов биосинтеза белка (происходящего в гранулярной ЭПС).
2. Синтез полисахаридов и превращение простых белков в гликопротеины.
3. Образование липонротеидов.
4. Формирование секреторных включений и выделение их из клетки (упаковка и секреция).
5. Образование первичных лизосом.
6. Формирование клеточных мембран.
7. Образование акросомы — структуры, содержащей ферменты, находящейся на переднем конце сперматозоида и необходимой для оплодотворения яйцеклетки, разрушения ее оболочек.

|
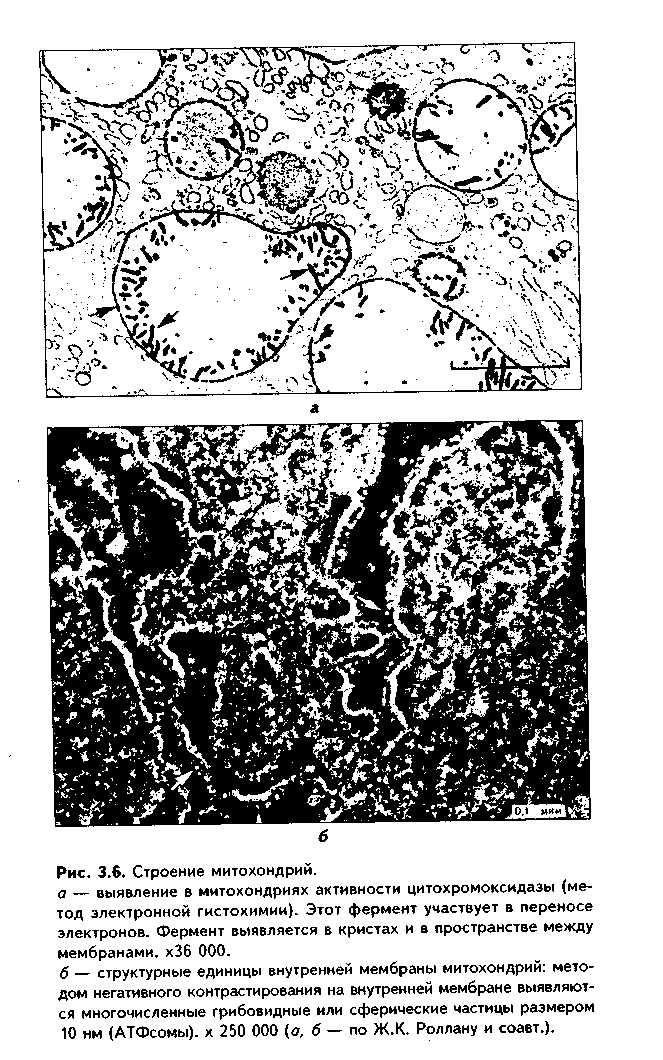
МИТОХОНДРИИ. Эти органеллы обеспечивают окисление органических соединений и синтез АТФ. Были открыты в 1890 году немецким ученым Р. Альтманом при помощи предложенного им метода окрашивания кислым фуксином, при этом выглядели в виде нитей и зерен (отсюда их название). Интересно, что сам Р. Альтман считал митохондрии бактериями, внедрившимися в клетку с паразитической целью, а в последующем ставшими симбионтами (симбиотическая теория происхождения митохондрий в настоящее время поддерживается многими исследователями).
Размеры митохондрий составляют от 0,5 до 7 мкм, а их общее число в клетке — от 50 до 5000. Эти органеллы хорошо видны в световом микроскопе, однако информация об их строении, получаемая при этом, скудна (рис. 3.5, а). Электронный микроскоп показал, что митохондрии состоят из двух мембран — наружной и внутренней, каждая из которых имеет толщину 7 нм (рис. 3.5, б, в, 3.6, а). Между наружной и внутренней мембранами имеется щель размером до 20 нм.
|
|
Внутренняя мембрана неровная, образует много складок, или крист. Эти кристы идут перпендикулярно поверхности митохондрии. На поверхности крист имеются грибовидные образования (оксисомы, АТФсомы или F,-частицы), представляющие собой АТФ-синтетазный комплекс (рис. 3.6) Внутренняя мембрана отграничивает матрикс митохондрии. Он содержит многочисленные ферменты для окисления пирувата и жирных кислот, а также ферменты цикла Кребса. Кроме того, в матриксе находятся мито-хондриальная ДНК, митохондриальные рибосомы, т-РНК и ферменты активации митохондриального генома. Внутренняя мембрана содержит белки трех типов: ферменты, катализирующие окислительные реакции; АТФ-син-тезатный комплекс, синтезирующий в матриксе АТФ; транспортные белки. Наружная мембрана содержит ферменты, превращающие липиды в реакционные соединения, участвующие затем в метаболических процессах матрикса. Межмембранное пространство содержит ферменты, необходимые для окислительного фосфорилирования. Т.к. митохондрии имеют свой геном, то они обладают автономной системой синтеза белка и могут частично строить собственные белки мембран.
Функции.
1. Обеспечение клетки энергией в виде АТФ.
2. Участие в биосинтезе стероидных гормонов (некоторые звенья биосинтеза этих гормонов протекают в митохондриях). Клетки-продуценты сте
роидных гормонов имеют крупные митохондрии со сложными крупными трубчатыми кристами.
3. Депонирование кальция.
4. Участие в синтезе нуклеиновых кислот. В некоторых случаях в результате мутаций митохондриалыюй ДНК возникают так называемые митохондриальные болезни, проявляющиеся широкой и тяжелой симптоматикой. ЛИЗОСОМЫ. Это мембранные органеллы, не видимые в световом микроскопе. Были открыты в 1955 году К. де Дювом при помощи электронного микроскопа (рис. 3.7). Представляют собой мембранные пузырьки, содержащие гидролитические ферменты: кислую фосфатазу, липазу, протеазы, нуклеазы и др., всего более 50 ферментов. Различают лизосомы 5 типов:
1. Первичные лизосомы, только что отделившиеся от транс-поверхности комплекса Гольджи.
2. Вторичные лизосомы, или фаголизосомы. Это лизосомы, которые соединились с фагосомой — фагоцитированной частицей, окруженной мембраной.
3. Остаточные тельца — это слоистые образования, формирующиеся в том случае, если процесс расщепления фагоцитированных частиц прошел не до конца. Примером остаточных телец могут быть липофусциновые включения, которые появляются в некоторых клетках при их старении, содержат эндогенный пигмент липофусцин.
4. Первичные лизосомы могут сливаться с погибающими и старыми органеллами, которые они разрушают. Такие лизосомы называются ауто-фагосомами.
5. Мультивезикулярные тельца. Представляют собой крупную вакуоль, в которой, в свою очередь, находятся несколько так называемых внутренних пузырьков. Внутренние пузырьки, очевидно, образуются путем отпочковывания внутрь от мембраны вакуоли. Внутренние пузырьки могут постепенно растворяться содержащимися в матриксе тельца ферментами.
Функции лизосом: 1. Внутриклеточное пищеварение. 2. Участие в фагоцитозе. 3. Участие в митозе — разрушении ядерной оболочки. 4. Участие во внутриклеточной регенерации.5. Участие в аутолизс — саморазрушении клетки после ее гибели.
Существует большая группа болезней, называемых лизосомными болезнями, или болезнями накопления. Они являются наследственными болезнями, проявляются дефицитом определенного лизосомального пигмента. При этом в цитоплазме клетки накапливаются непереваренные продукты

|
обмена веществ (гликоген, гликолиниды, белки, рис. 3.7, б,в), что ведет к постепенной гибели клетки. ПЕРОКСИСОМЫ. Пероксисомы — это оргаиеллы, напоминающие ли-зосомы, но содержащие ферменты, необходимые для синтеза и разрушения эндогенных перекисей — нероксидазу, каталазу и другие, всего до 15. В электронном микроскопе представляют сферические или эллипсоидные пузырьки с умеренно плотной сердцевиной (рис. 3.8). Образуются пероксисомы путем отделения пузырьков от гладкой ЭПС. В эти пузырьки затем миг-рируют ферменты, которые синтезируются отдельно в цитозоле или в гранулярной ЭПС
Функции пероксисом: 1. Являются, наряду с митохондриями, органеллами утилизации кислорода. В результате в них образуется сильный окислитель Н202. 2. Расщепление при помощи фермента каталазы избытка перекисей и, таким образом, защита клеток от гибели. 3. Расщепление при помощи синтезируемых в самих пероксисомах перекисей токсических продуктов, имеющих экзогенное происхождение (детоксикация). Такую функцию выполняют, например, пероксисомы печеночных клеток, клеток почек. 4. Участие в метаболизме клетки: ферменты пероксисом катализируют расщепление жирных кислот, участвуют в обмене аминокислот и других веществ.

Существуют так называемые пероксисомные болезни, связанные с дефектами ферментов пероксисом и характеризующиеся тяжелыми поражениями органов, что ведет к смерти в детском возрасте. НЕМЕМБРАННЫЕ ОРГАНЕЛЛЫ
РИБОСОМЫ. Это оргаиеллы биосинтеза белка. Они состоят из двух рибонуклеоиротеидных субьединиц — большой и малой. Эти субъединицы могут соединяться вместе, при этом между ними располагается молекула информационной РНК. Есть свободные рибосомы — рибосомы, не связанные с ЭПС. Они могут быть одиночными и в виде полисом, когда на одной молекуле и-РНК находятся несколько рибосом (рис. 3.9). Вторая разновидность рибосом — связанные рибосомы, прикрепленные к ЭПС.

|
Функция рибосом. Свободные рибосомы и полисомы осуществляют биосинтез белка для собственных потребностей клетки.

Связанные на ЭПС рибосомы синтезируют белок на "экспорт", для нужд всего организма (например, в секреторных клетках, нейронах и др.).
МИКРОТРУБОЧКИ. Микротрубочки являются органеллами фибриллярного типа. Они имеют диаметр 24 им и длину до нескольких мкм. Это прямые длинные полые цилиндры, построенные из 13 периферических нитей, или протофиламентов. Каждая нить образована глобулярным белком тубулином, который существует в виде двух субъединиц — аир (рис. 3.10). В каждой нити эти субъединицы располагаются поочередно. Нити в микротрубочке имеют спиральный ход. В стороны от микротрубочек отходят ассоциированные с ними молекулы белков (ассоциированные с микротрубочками протеины, или MAP). Эти белки стабилизируют микротрубочки, а также связывают их с другими элементами цитоскелета и орга-неллами. С микротрубочками связан также белок кииезин, который представляет собой фермент, расщепляющий АТФ и преобразующий энергию ее распада в механическую энергию. Одним концом кииезин связывается с определенной органеллой, а другим за счет энергии АТФ скользит вдоль микротрубочки, перемещая таким образом органеллы в цитоплазме

|
Микротрубочки являются очень динамичными структурами. Они имеют два конца: (-) и (+) — концы. Отрицательный конец является местом деполимеризации микротрубочки, тогда как на положительном конце происходит их наращивание за счет новых молекул тубулина. В некоторых случаях (базальное тельце) отрицательный конец как бы заякоривается, и распад здесь прекращается. В результате происходит увеличение размеров ресничек из-за наращивания на (+) — конце.
Функции микротрубочек заключаются в следующем. 1. Выполняют роль цитоскелета;
2. Участвуют в транспорте веществ и органелл в клетке;
3. Участвуют в образовании веретена деления и обеспечивают расхождение хромосом в митозе;
4. Входят в состав центриолей, ресничек, жгутиков.
Если клетки обработать колхицином, разрушающим микротрубочки цитоскелета, то клетки изменяют свою форму, сжимаются, теряют способность к делению.
МИКРОФИЛАМЕНТЫ. Это второй компонент цитоскелета. Есть два вида микрофиламентов: 1) актиновые; 2) промежуточные. Кроме того, цитоскелет включает множество вспомогательных белков, которые связывают филаменты друг с другом или с другими клеточными структурами.
Актиновые филаменты построены из белка актина и образуются в результате его полимеризации. Актин в клетке находится в двух формах: 1) в растворенной форме (G-актин, или глобулярный актин); 2) в полимери-зованиой форме, т.е. в виде филаментов (F-актин). В клетке существует динамическое равновесие между 2 формами актина. Как и в микротрубочках, в актиновых филаментах имеются (+) и (-) — полюсы, и в клетке идет постоянный процесс распада этих филамент на отрицательном и созидание на положительном полюсах. Этот процесс называется тредмил-лингом. Он играет важную роль в изменении агрегатного состояния цитоплазмы, обеспечивает подвижность клетки, участвует в перемещении ее органелл, в формировании и исчезновении псевдоподий, микроворсинок, протекании эндоцитоза и экзоцитоза. Микротрубочки создают каркас микроворсинок, а также участвуют в организации межклеточных включений.
Промежуточные филаменты — филаменты, имеющие толщину, большую, чем у актиновых филаментов, но меньшую, чем у микротрубочек. Это самые стабильные филаменты клеток. Выполняют опорную функцию. Например, эти структуры лежат по всей длине отростков нервных клеток, в области десмосом, в цитоплазме гладких миоцитов. В клетках разного типа промежуточные филаменты отличаются по составу. В нейронах образуются ней-рофиламенты, состоящие из трех различных полипентидов. В клетках нейроглии промежуточные филаменты содержат кислый глиальный белок. В эпителиальных клетках содержатся кератиновые филаменты (тонофила-менты) (рис. 3.11).

|
В мышечных клетках (за исключением миоцитов сосудов) промежуточные филаменты состоят из белка десмина. В различных клетках мезенхимного происхождения, в том числе и в миоцитах сосудов, содержатся виментиновые филаменты.
|
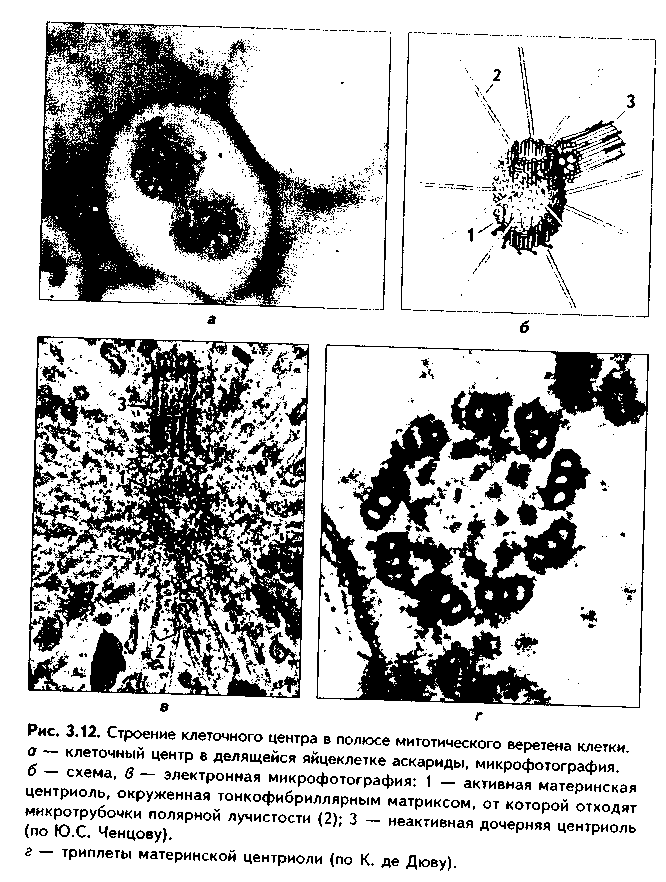
КЛЕТОЧНЫЙ ЦЕНТР (рис. 3.12). Это видимая и световой микроскоп органелла, однако ее тонкое строение позволил изучить только электронный микроскоп. В интерфазной клетке клеточный центр состоит из двух цилиндрических полостных структур длиной до 0,5 мкм и диаметром до 0,2 мкм. Эти структуры называются центриолями. Они образуют диплосому. В диплосоме дочерние центриоли лежат под прямым углом друг к другу. Каждая центриоль состоит из расположенных по окружности 9 триплетов микротрубочек, которые частично сливаются по длине. Кроме микротрубочек, в состав цептриолей входят "ручки" из белка динеина, которые соединяют соседние триплеты в виде мостиков. Центральные микротрубочки отсутствуют, и формула центриолей — (9хЗ)+0. Каждый триплет микротрубочек связан также со структурами сферической формы — сателлитами. От сателлитов расходятся в стороны микротрубочки, образуя центросферу.
Центриоли являются динамичными структурами и претерпевают изменения в митотическом цикле. В неделящейся клетке парные центриоли (центросома) лежат в околоядерной зоне клетки. В S-периоде митотического цикла они дуплицируются, при этом под прямым углом к каждой зрелой центриоли образуется дочерняя центриоль. В дочерних центриолях вначале имеется только 9 единичных микротрубочек, но по мере созревания центриолей они превращаются в триплеты. Далее пары центриолей расходятся к полюсам клетки, становясь центрами организации микротрубочек веретена деления.
Значение центриолей.
1. Являются центром организации микротрубочек веретена деления.
2. Образование ресничек и жгутиков.
3. Обеспечение внутриклеточного передвижения органелл. Некоторые авторы считают, что определяющими функциями клеточного
центра являются вторая и третья функции, поскольку в растительных клетках центриоли отсутствуют, тем не менее и в них образуется веретено деления.
РЕСНИЧКИ И ЖГУТИКИ (рис. 3.13). Это специальные органеллы движения. Они имеются в некоторых клетках — сперматозоидах, эпителиоцитах трахеи и бронхов, семявыводящих путей мужчины и др. В световом микроскопе реснички и жгутики выглядят как тонкие выросты. В электронном микроскопе установлено, что в основании ресничек и жгутиков лежат мелкие гранулы — базальные тельца, одинаковые по строению с центриолями. От базального тельца, являющегося матрицей при росте ресничек и жгутиков, отходит тонкий цилиндр из микротрубочек — осевая нить, или аксонема. Она состоит из 9 дуплетов микротрубочек, на которых находятся "ручки" из белка динеина. Аксонема покрыта цитолеммой. В центре находится пара микротрубочек, окруженная специальной оболочкой — муфтой, или внутренней капсулой. От дуплетов к центральной муфте идут радиальные спицы. Следовательно, формула ресничек и жгутиков — (9х2)+2.
Основу микротрубочек жгутиков и ресничек составляет несократимый белок тубулин. Белок "ручек" — динеин — обладает АТФазной активное -гыо: расщепляет АТФ, за счет энергии которой происходит смещение дуплетов микротрубочек друг по отношению к другу. Так совершаются волнооб-ралные движения ресничек и жгутиков.
Существует генетически обусловленное заболевание — синдром Карта-гснера, при котором в аксонеме отсутствуют либо динеиновые ручки, либо центральная капсула и центральные микротрубочки (синдром неподвижных ресничек). Такие больные страдают рецидивирующими бронхитами, синуситами и трахеитами. У мужчин из-за неподвижности спермиев отмечается бесплодие.
МИОФИБРИЛЛЫ находятся в мышечных клетках и миосимпластах, и их строение рассматривается в теме "Мышечные ткани". Нейрофибрил-лы находятся в нейронах и состоят из нейротубул и нейрофиламентов. Их функция — опорная и транспортная.
ВКЛЮЧЕНИЯ
Включения — это непостоянные компоненты клетки, не имеющие строго постоянной структуры (их структура может меняться). Выявляются в клетке только в определенные периоды жизнедеятельности или жизненного цикла.

|
КЛАССИФИКАЦИЯ ВКЛЮЧЕНИЙ.
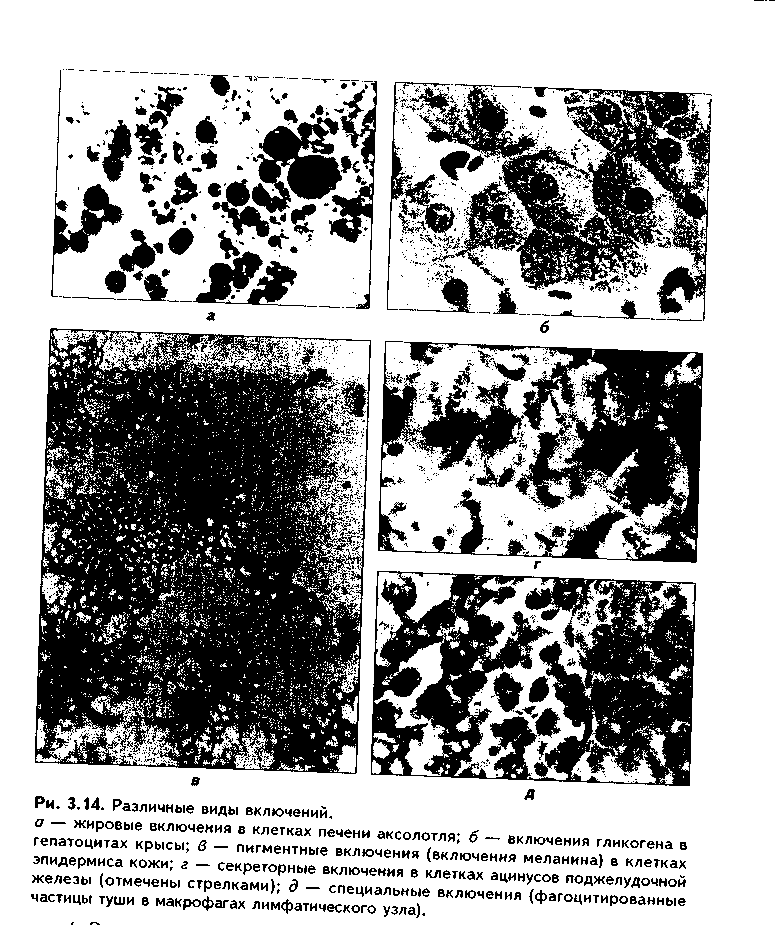
1. Трофические включения представляют собой депонированные питательные вещества. К таким включениям относятся, например, включения гликогена, жира.
2. Пигментные включения. Примером таких включений являются гемоглобин в эритроцитах, меланин в меланоцитах. В некоторых клетках (нервные, печеночные, кардиомиоциты) при старении в лизосомах накапливается пигмент старения коричневого цвета липофусцин, не несущий, как полагают, определенной функции и образующийся в результате изнашивания клеточных структур. Следовательно, пигментные включения представляют собой химически, структурно и функционально неоднородную группу. Гемоглобин участвует в транспорте газов, меланин выполняет защитную функцию, а липофусцин является конечным продуктом обмена. Пигментные включения, за исключением лииофусциновых, не окружены мембраной.
3. Секреторные включения выявляются в секреторных клетках и состоят из продуктов, представляющих собой биологически активные вещества и другие необходимые для осуществления функций организма вещества (включения белка, в том числе и ферментов, слизистые включения в бокаловидных клетках и др.). Эти включения имеют вид окруженных мембраной пузырьков, в которых секретируемый продукт может иметь различную электронную плотность и часто окружен светлым бесструктурным ободком. 4. Экскреторные включения — включения, подлежащие выведению из клетки, поскольку состоят из конечных продуктов обмена. Примером являются включения мочевины в клетках почки и т.д. По структуре похожи на секреторные включения.
5. Специальные включения — фагоцитированные частицы (фагосо-мы), поступающие в клетку путем эндоцитоза (см. ниже). Различные виды включений представлены на рис. 3.14.
|
|
|

Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...

Поперечные профили набережных и береговой полосы: На городских территориях берегоукрепление проектируют с учетом технических и экономических требований, но особое значение придают эстетическим...

История развития хранилищ для нефти: Первые склады нефти появились в XVII веке. Они представляли собой землянные ямы-амбара глубиной 4…5 м...

Типы сооружений для обработки осадков: Септиками называются сооружения, в которых одновременно происходят осветление сточной жидкости...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!